Chromatin Compaction as a Topological Problem
Presenter
September 20, 2007
Keywords:
- DNA sequences
MSC:
- 92D20
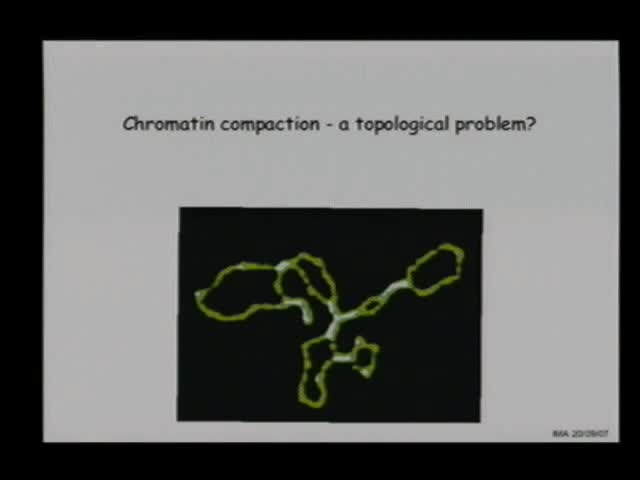
Abstract
In both bacteria and eukaryotes, maintaining DNA compaction is
a sine qua non of chromatin function. At the same time
accessibility to transcribing, replicating and recombining
enzymes must be maintained. I will argue that these twin
requirements can be viewed in the context of the overall
topology of, for bacteria, the DNA itself, and of, for
eukaryotes, the 30 nm chromatin fibre. In particular, the
plectonemic form of supercoiled ropes, either DNA or 30 nm
fibres, must, in principle, contain distinguishable structures
(interwindings, apical loops, branch points, hammerheads) in
contrast to a more monotonous toroid.
In exponentially growing Escherichia coli the 2-start helical
interwindings of the plectonemic form of plasmid DNA are
stabilised by different nucleoid-associated proteins (NAPs).
Nucleation of binding of the NAP H-NS at high affinity sites
results in gene silencing and plectoneme stabilisation.
Plectonemes are also recognised by RNA polymerase and certain
recombinases. The binding of RNA polymerase to loop structures
formed at some promoters facilitates binding and for the
tyrT
promoter potentially locates the thermally unstable -10 hexamer
adjacent to the interwindings of a negatively supercoiled
plectoneme.
In eukaryotes the requirement for DNA compaction is greater
than in bacteria. Whereas a simple DNA plectoneme compacts DNA
by ~~2.5 fold, compaction factors of up to 10000 are required
in the eukaryotic nucleus. The initial mechanism for compacting
DNA is the tight wrapping of ~146 bp in a nucleosome core
particle, resulting in a compaction of ~9-10-fold. In vivo
these particles can be accurately positioned such that the
midpoint of the bound DNA is approximately defined. We have
derived from accurately mapped in vivo positions in yeast a
translational positioning signal that identifies the midpoint
of histone octamer-bound DNA. The minimal signature is 75% of reported and also our newly
determined mapped positions in yeast. This putative positioning
signal occurs on average once every ~60 bp in yeast genomic DNA
sequences. From this apparent redundancy we infer that the
preferred positioning of nucleosomes in an array requires an
'organiser' to select a nucleosome for nucleating an array.
This organiser could be a strong intrinsic DNA positioning
signal or a transcription factor. We present evidence that the
DNA sequences specifying 5' proximal (-1 position) nucleosomes
of several genes can, under more physiological conditions,
outcompete in vitro the strong 601 positioning sequence
originally selected by salt gradient dialysis in vitro.
The next stage in the compaction of eukaryotic chromatin is the
folding of a nucleosome array into a '30 nm' fibre. We have
calculated the dependence of the diameter and packing density
of chromatin fibres on linker length and conclude that all
current measurements are consistent with a model in which at
short linker lengths (corresponding to a nucleosome repeat
length of ~ 177 bp) the linker histone can supercoil a 2-start
crossed-linker fibre into a helical-ribbon form by changing the
exit and entry trajectories of DNA. As the linker length
increases in increments of 10 bp (< the 10.5 bp helical repeat
of DNA) at a certain point the fibre relaxes into a
crossed-linker form with a higher packing density. On this
model the 30 nm fibre has a variable topology but maintains a
constant packing of nucleosomes.
References:
Maurer, S., Fritz., J., Muskhelishvili, G. and Travers, A. RNA
polymerase and an activator form discrete subcomplexes in a
transcription initiation complex. EMBO J. 25, 3784-3790 (2006).
Travers, A. and Muskhelishvili, G. A common topology for
bacterial and eukaryotic transcription initiation? EMBO Rep. 8,
147-151 (2007).
Bouffartigues, E., Buckle, M., Baudaut, C., Travers, A. and
Rimsky, S. High affinity sites direct the cooperative binding
of H-NS to a regulatory element required for transcriptional
silencing. Nat. Struct. Mol. Biol. 14, 441-448 (2007).
Lang, B. et al. High affinity DNA binding sites for H-NS
provide a molecular basis for selective silencing within
proteobacterial genomes. Submitted for publication (2007).
Wu, C., Bassett, A. and Travers, A. A variable topology for the
'30 nm' chromatin fibre. Submitted for publication (2007).
Collaborators:
MRC-LMB, Cambridge: M. Madan Babu, Mark Churcher, Edwige
Hiriart, Benjamin Lang, Chenyi Wu
DAMTP, University of Cambridge: Graeme Mitchison
ENS, Cachan: Cyril Baudaut, Emeline Bouffartigues, Malcolm
Buckle, Sylvie Rimsky
Università di Roma "La Sapienza": Eleonora Agricola, Micaela
Caserta, Ernesto Di Mauro, Leonora Verdone
Università di Camerino: Claudio Gualerzi, Cynthia Pon, Stefano
Stella
Jacobs University, Bremen: Claudia Burau, Nicolas Blot, Jurgen
Fritz, Marcel Geertz, Sebastian Maurer, Ramesh Mavathur, Georgi
Muskhelishvili